|
|
Главная - Статьи - Гистологическая структура ресничного ганглия...
Е. О. Юдин
Гистоструктура ресничного ганглия и морфометрический анализ нейроцитов у представителей отряда Хищные.
Введение
Ресничный ганглий (РГ) у домашних животных, на наш взгляд, является наиболее изученным из всех автономных ганглиев головы. Обстоятельно изучены его форма, внешнее строение, топография и связи с ветвями глазодвигательного и тройничного нервов как у человека [4, 11], так и у птиц и млекопитающих [2, 6, 8, 9, 10, 12], в том числе и у пушных зверей клеточного содержания. Несмотря на достаточную степень изученности топографии и связей РГ, в научной литературе все еще остается дискуссионным вопрос об участии РГ в симпатической иннервации структур глазного яблока. Многие авторы выска-зывают традиционную точку зрения о наличии у РГ млекопитающих симпатического корня [9, 11]. В то же время в более поздних работах наличие подобной анатомической связи отрицается [2, 4, 6, 12], в том числе и гистохимическими методами [7].
Достаточно хорошо изучено и гистологическое строение РГ. Так, в работе Е. Н. Соловьевой [3] мы находим сведения о морфологии нейроцитов РГ у человека, кошки и собаки. Приведены краткие морфометрические данные. Подробные сведения о гистологическом строении РГ макаки, в т.ч. данные об ультраструктуре его нейроцитов содержатся в работе Y. L. Zhang et al. [12]. Авторы приводят достаточно обстоятельные морфометрические данные и проводят сравнительный морфометрический анализ нейроцитов РГ макаки с таковыми у других млекопитающих. D. A. Johnson и D. Purves [6] изучали гистологическое строение ресничного ганглия кролика в онтогенетическом аспекте.
Вместе с тем в доступных нам специальных источниках мы не
нашли сведений о гистологическом строении РГ у пушных зверей клеточного
содержания, а также данных сравнительного морфометрического анализа нейроцитов
РГ у хищных млекопитающих.
В связи с вышеизложенным, нами была поставлена цель – изучить гистологическую
структуру РГ у собаки домашней, лисицы обыкновенной, норки, соболя и кошки
домашней и провести сравнительный морфометрический анализ нейроцитов
РГ у этих животных.
Материал и методы исследования
В качестве объекта исследования были использованы трупы пушных зверей и домашних животных, относящихся к классу млекопитающих из отряда хищных: семейство Собачьи (лисица обыкновенная, собака домашняя), Куньи (норка, соболь) и Кошачьи (кошка домашняя). Материал исследования – ресничный ганглий, полученный в течение 6–10 часов после убоя животных, фиксировали в 5–7% растворе нейтрального формальдегида, после чего уплотняли заливкой в парафин по общепринятой методике. Срезы толщиной 5–8 мкм получали на санном микротоме.
Для изучения общей гистологической картины срезы окрашивали гематоксилином Вейгерта и спиртовым раствором эозина. Соединительнотканные волокна выявляли по методу Ван Гизон. Базофильное вещество в нейроцитах изучали на срезах, окрашенных тионином по Нисслю. Изготовленные препараты изучали под световым микроскопом «BIOLAR» и фотографировали цифровой камерой «OLYMPUS». Полученные цифровые изображения подвергались морфометрической обработке при помощи разработанной нами программы ScreenMeter [5]. Измерялись больший (длина) и меньший (ширина) диаметры тела нейроцита, а также больший и меньший диаметры его ядра, после чего рассчитывались по соответствующим формулам [1] площади клетки и ее ядра, эксцентриситет (степень вытянутости) и ядерно-цитоплазменное отношение (ЯЦО). Полученный цифровой материал подвергался статистической обработке с использованием пакета анализа STATISTICA 6.0. По полученным выборкам рассчитывались описательные статистики, после чего имеющиеся данные проверялись на соответствие их закону о нормальном распределении при помощи критерия Шапиро-Уилка. В сравниваемых выборках оценивалась степень гомогенности их дисперсий методом Брауна-Форсайта. В случае если данные не соответствовали закону о нормальном распределении величин, или дисперсии выборок достоверно отличались, для подтверждения отличий между выборками применялся непараметрический критерий Манна-Уитни. Если же сравниваемые выборки отвечали обоим условиям (нормальность распределения и гомогенность дисперсий), то использовался t-тест по Стъюденту. Межвидовые сравнения производились при помощи непараметрического критерия Краскелла-Уоллиса. Для обнаружения корреляционных зависимостей между исследуемыми параметрами использовался метод ранговой корреляции по Спирману.
Результаты собственных исследований
В результате проведенных исследований было установлено, что у изученных животных ресничный ганглий представляет собой компактное образование, имеющее на срезе овальную или овально-изогнутую (бобовидную), реже округлую форму. У норки РГ чаще имеет на срезе ромбовидную или многоугольную форму. Снаружи ганглий покрыт соединительнотканной капсулой, образованной плотно лежащими коллагеновыми волокнами, между которыми размещаются ядра фиброцитов и фибробластов веретеновидной формы. Капсула имеет неравномерную толщину и, как правило, утолщается в тех местах, где происходит вступление в РГ преганглионарных волокон, либо выхождение его постганглионарных ветвей – коротких ресничных нервов. От капсулы вглубь ганглия отходят соединительнотканные прослойки, которые, разветвляясь, принимают участие в формировании соединительнотканной капсулы нейроцитов ганглия.
Основная в функциональном отношении ткань РГ образована телами нервных клеток и нервными волокнами. Нами отмечено два основных типа строения РГ. Чаще всего тела нейроцитов располагаются в ганглии равномерно, без видимого разбиения на группы или ядра, а в промежутках между ними находятся пучки нервных волокон, срезанных в различных плоскостях. Несколько реже наблюдается другой вариант строения, когда тела нервных клеток сосредоточены 1–2 группами у полюсов РГ, а центральная часть узла заполнена волокнистым компонентом нервной ткани.
Тела нейроцитов, формирующих РГ, имеют эллипсоидную, реже округлую форму. На срезах, окрашенных гематоксилином и эозином, цитоплазма клеток имеет светло-розовую окраску с легкой базофильной зернистостью. Ядра клеток крупные, светлые, правильной эллипсоидной или округлой формы. Ядерная мембрана четко очерчена. Ядра содержат одно или два (очень редко) интенсивно окрашенных округлых ядрышка. Помимо ядрышка в ядре обнаруживаются глыбки хроматина различного размера и неправильной формы, хаотично расположенные в ядре (рис. 1). На срезах, окрашенных тионином по Нисслю, в цитоплазме нервной клетки наблюдается равномерно распределенная базофильная зернистость. Иногда базофильное вещество может быть в большей степени сконцентрировано вокруг ядра клетки (рис. 2).

Рис. 1. Взаимоотношения нейроцита с глиоцитами в ресничном
ганглии у собаки.
Окраска гематоксилином Вейгерта и эозином. Об. х100, ок. х12,5
1 – тело нейроцита; 2 – ядро; 3 – ядрышко; 4 – ядра глиоцитов.
Снаружи тела нейроцитов окружены двухслойной капсулой. Внутренний ее слой, непосредственно прилегающий к телу клетки, сформирован одним рядом ганглионарных глиоцитов (рис. 1). На препаратах нередко встречаются участки, где два соседних нейроцита тесно прилегают друг к другу. В таком случае глиоциты в месте их контакта отсутствуют. На срезах, окрашенных гематоксилином и эозином, цитоплазма глиоцитов имеет светло-розовую окраску. Ядра их мелкие, интенсивно окрашены. Ядра содержат по одному мелкому ядрышку. Глиоциты тесно прилегают к телу нейроцита. Иногда на теле нейроцита заметно небольшое вдавление, в котором располагается глиоцит.
Наружный слой капсулы нейроцита сформирован соединительнотканными волокнами, между которыми рассредоточены ядра фиброцитов и фибробластов, имеющие эллипсоидную форму.
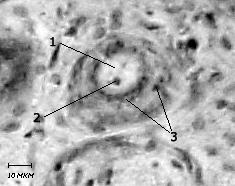
Рис. 2. Базофильное вещество в цитоплазме нейроцита ресничного
ганглия кошки.
Окраска тионином по Нисслю. Об. х20, ок. х12,5.
1 – ядро клетки, 2 – ядрышко, 3 – глыбки базофильного вещества.
Рассматривая гистологическое строение РГ у изученных видов
животных можно прийти к выводу о том, что РГ у них построен однотипно
и имеет строение, в целом характерное для большинства экстраорганных автономных
ганглиев. Для выявления видовых отличий нами был применен
комплекс морфометрических методов.
В табл. 1 сведены данные о средних значениях изученных нами параметров
нейроцитов РГ. Цифры приведены в формате M±m, где M –
среднее арифметическое выборки, m – стандартная ошибка среднего.
Табл. 1 Средние значения и ошибки среднего по исследованным параметрам нейроцитов РГ
| Параметр | Собака | Лисица | Норка | Соболь | Кошка |
| Длина клетки, мкм. | 40,4±0,36 |
42,4±0,29 |
30,8±0,46 |
33,3±0,34 |
43,3±0,69 |
| Ширина клетки, мкм. | 28,8±0,3 |
31,9±0,24 |
19,9±0,46 |
24,7±0,24 |
33,9±0,55 |
| Площадь клетки, мкм. кв. | 930±16,1 |
1081±14 |
498±16 |
675±13 |
1194±37 |
| Эксцентриситет клетки | 0,7±0,008 |
0,6±0,006 |
0,73±0,01 |
0,6±0,006 |
0,6±0,01 |
| Длина ядра, мкм. | 15,4±0,12 |
16,2±0,08 |
11,7±0,13 |
12,7±0,06 |
18,0±0,23 |
| Ширина ядра, мкм. | 12,2±0,12 |
13,5±0,07 |
8,5±0,21 |
10,2±0,06 |
14,4±0,17 |
| Площадь ядра, мкм. кв. | 150,6±2,6 |
173,5±1,7 |
79,1±2,3 |
103,3±0,9 |
207,9±4,8 |
| Эксцентриситет ядра | 0,6±0,007 |
0,5±0,006 |
0,65±0,01 |
0,6±0,006 |
0,6±0,01 |
| ЯЦО | 0,2±0,004 | 0,2±0,003 | 0,22±0,01 | 0,2±0,03 | 0,2±0,01 |
Межвидовые сравнения по критерию Краскелла-Уоллиса подтверждают наличие достоверных отличий по всем сравниваемым выборкам (рис. 3). Линейные параметры (длина и ширина) ганглионарных нейроцитов и их ядер возрастают в ряду: норка – соболь – собака – лисица – кошка.

Рис.3. Размеры нейроцитов и их ядер у изученных видов
животных.
А – длина клетки, Б – ширина клетки, В – длина ядра, Г – ширина ядра,
1 – лисица, 2 – кошка, 3 – собака, 4 – норка, 5 – соболь.
Изучение корреляционных зависимостей показывает наличие достоверных связей между изучаемыми параметрами. Наибольшего внимания, на наш взгляд, заслуживают взаимоотношения между размерами нейроцитов и эксцентриситетом клетки (рис. 4). Как показал корреляционный анализ, между площадью клетки и ее эксцентриситетом имеется слабая отрицательная связь, т. е. с увеличением среднего значения площади клеток РГ, форма их становится более близка к округлой, нежели к эллипсоидной.
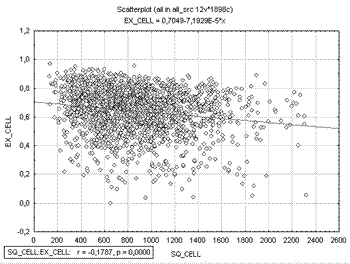
Рис.4. Диаграмма рассеяния, отражающая корреляционные взаимоотношения между пло-щадью клетки (по горизонтали) и ее эксцентриситетом (по вертикали).
Обсуждение
Анализ результатов собственных исследований и их сопоставление с данными литературы показывают, что гистологическое строение РГ у исследованных животных аналогично таковому у других млекопитающих. Так, результаты наших исследований согласуются со сведениями об общем гистологическом строении РГ и его нейроцитов у человека, кошки и собаки [3], кролика [6] и макаки [12]. Мы согласны с мнением Y. L. Zhang, C. K. Tan and W. C. Wong [12] о том, что нейроциты РГ у млекопитающих морфометрически можно отнести к одной нейрональной популяции. У изученных нами животных мы также наблюдали унимодальное распределение по исследованным морфометрическим параметрам нейроцитов. Последнее является фактором, значительно отличающим морфологию нервных клеток РГ млекопитающих от таковых у птиц. Авторы, исследовавшие РГ птиц, отмечают две популяции нейроцитов: более крупные, называемые ресничными клетками и более мелкие хороидальные нейроциты [8, 10].
Результаты наших исследований показывают наличие достоверных межвидовых отличий по всем изученным морфометрическим параметрам нейроцитов РГ. Y. L. Zhang, C. K. Tan and W. C. Wong считают, что размеры нейроцитов РГ и их ядер коррелируют с размерами животного. На наш взгляд, подобная взаимосвязь присутствуют, однако мы считаем, что размеры животного являются не единственным фактором, влияющим на морфометрические параметры нервных клеток РГ. Так, показатели нейроцитов РГ кошки и лисицы преобладали над таковыми у собаки (рис. 3), несмотря на то, что собака является более крупным животным, нежели кошка и лисица. Общеизвестно, что зрачок кошки и лисицы, в отличие от собаки может изменять не только свои размеры, но и форму – от крупного округлого отверстия до узкой вертикальной щели. Мы предполагаем, что более сложное строение цилиарной мышцы у этих животных обусловливает более крупные размеры нейроцитов РГ.
Выводы
Таким образом, проведенные нами исследования позволяют
утверждать, что РГ у изученных животных имеет типичное строение, свойственное
автономным ганглиям, и представляет собой компактную
группу нейроцитов. При проведении морфометрического анализа
выявлены достоверные (p<0,05) межвидовые отличия по всем исследованным
параметрам. Выявленные видовые отличия, на наш взгляд, наглядно характеризуют
степень развития зрачкового рефлекса у изученных видов животных, что в
свою очередь, безусловно связано с образом их жизни, способом добывания
пищи и условиями окружающей среды.
Список литературы
- Автандилов, Г. Г. Медицинская морфометрия / Г. Г. Автандилов. – М.: Медицина, 1990. – 384 с.: ил.
- Ефимов, С. И. Взаимоотношения глазничного нерва с ресничным ганглием у пуш-ных зверей / С. И. Ефимов // Экологические аспекты функциональной морфологии пуш-ных зверей. – Омск: изд-во ОмСХИ, 1989. – С. 15-18
- Соловьева, Е. Н. О возрастных изменениях и «окончатости» нервных клеток рес-ничного узла у человека, кошки и собаки / Е. Н. Соловьева // Вопросы нейроморфологии. – Ярославль: Ярославский мед. ин-т, 1959. – С. 81 – 92
- Цыбулькин, А. Г. Индивидуальная изменчивость внешнего строения и топографии ресничного узла человека / А. Г. Цыбулькин // Морфология. – 2003. ¬– № 6. – С. 34 – 37
- Юдин, Е. О. Разработка программного обеспечения для морфометрических иссле-дований / Ю. С. Гайдученко, Е. О. Юдин // Морфология. – 2006. – № 6. – С. 34
- D. A. Johnson, D. Purves Post-natal reduction of neural unit size in the rabbit ciliary ganglion. – Journal of the Physiology, №318. – 1981, P. 143-159.
- Immunohistochemical characterization of neurons in the porcine ciliary ganglion / Kaleczyc J, Juranek J, Calka J, Lakomy M. // Polish journal of veterinary sciences. – 2005. – №8. – P. 65 – 72.
- H. Oehme Das Ganglion ciliare der Rabenvogel (Corvidae) Anatomischer anzeiger Bd. 123, 1968, S. 261-277.
- E. Pastea Contributions a l’etude macroscopique des relations orthoparasympathiques craniennes chez le mouton. Anatomischer anzeiger, Bd 137, 1975, P. 492 – 503.
- M. Radzimirska The morphology, topography and cytoarchitectonics of the ciliary ganglion in the domestic turkey (Meleagris gallopavo domesticus). Folia Morphologica, 2003 Nov;62(4): P. 389-391.
- Z. Sinnreich, H. Nathan The ciliary ganglion in man (anatomical observations). – Anatomisher Anzeiger, 1981, №150(3), P. 287 – 297.
- Y. L. Zhang, C. K. Tan and W. C. Wong The ciliary ganglion of the monkey: a light and electron microscope study. Journal of Anatomy. - №184, 1994. – P. 251-260.
Реферат
Юдин, Е. О. Гистоструктура ресничного ганглия и морфометрический анализ нейроцитов у представителей отряда Хищные / Е. О. Юдин // Нерврологический вестник им. В. М. Бехтерева. - Казань, 2008. - Т. XL, вып. 4. - С.86-90.
| Подписка на рассылку новостей проекта: |